第三章 突 变
变幻中徘徊之物,固定于永恒的思想中。
——歌德
1.“跳跃式”的突变——自然选择的工作场地
前面我们论证的基因结构的持久性,提出了一般的论据,但仅有此并不具备强烈的说服力。“例外提供了法则的证明”,如果子女与父母之间的相似性没有例外的话,我们就不会潜心研究微观的遗传机制,更不会去揭示宏观的自然选择原理。
我把刚刚提到的自然选择作为介绍有关实验的导引。我们知道达尔文犯了一个巨大的错误,他把在最单一的群体中也会出现的细微、偶然的连续变异当做是自然选择的题中之义。这种错误的想法被后来的实验证实了,因为有些变异确实是不遗传的。打个比方,如果拿来一捆纯种的大麦,测量每一个麦穗的麦芒长度,然后根据这些数据作图,如图7所示。这是以麦穗数相对于麦芒的长度而作的图。中等长度占优势,长度增加和长度减少,麦穗数都要减少。把麦芒超过平均长度的麦穗拿出来(图中涂黑色的那一组),把它们的种子播种并等候其长出来。对新长出来的大麦作同样的统计,我们按照达尔文的理论将会预期一条极大值向右方移动的曲线。也就是说,他可以通过选择来增加麦芒的平均长度。但是如果选用的是绝对纯种的大麦,就不会是这种情况了。选择出来的大麦播种,经过统计作图,与第一
条曲线是一样的。如果选择麦芒特别短的做实验,进行统计作图后也是一样的。细微的、连续的变异是不遗传
的,因此选择没有效果。这些变化不是以遗传物质的结构为基础的,它们是偶然出现的。但是,在上面的讨论中,我们忽视了突变。40多年前,荷兰人德佛里斯(de Vries)发现,在完全纯种繁殖的后代中也有极少数的例外个体,比如有几万分之二三的概率,它们身上出现了细微但却是“跳跃式”的变化。这种“跳跃式”的变化不一定非常大,但却是一种不连续的变化——在没有改变和改变之间没有过渡形式,所以德佛里斯称之为突变。突变的理论容易使一个物理学家想起量子论——在两个相邻的能量级之间没有中间能量,因此他愿意把德佛里斯的突变理论称为生物学的量子论。事实上,突变是由基因分子中的量子跃迁而引起的。1902年,当德佛里斯第一次发表他的理论观点时,量子论问世仅有两年时间。因此,由另一代学者去发现两者之间的关系也就是很正常的了。
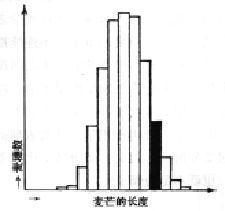
图7 纯种大麦的麦芒长度的统计。
涂黑色的那组是选做播种的(
本图细节并不是根据实际画出的,
仅作说明之用)
2.它们生育同样的后代,即它们是完全地遗传了下来
就像一直没有改变的特性一样,突变也是可以完全地遗传下去的。例如,上面提到的大麦,在它的首次收获中可以找到一些麦穗的麦芒,而且会超过图7所展示的变异范围,完全没有麦芒就是一个例子。这就是德佛里斯所说的突变,它们的后代将都没有麦芒,完全一样。
因此,德佛里斯的突变一定是遗传学机制中的一种变化,它必须也只能由遗传学的变化来作出科学合理的解释。实际上,繁育试验给我们展示了遗传机制的重要秘密,它采取的方法就是研究分析杂交后获得的后代,这些后代是由已经突变的个体和没有突变的个体或具有多重突变的个体杂交而来。另一方面,达尔文曾经论述到的自然选择的原料,在繁育过程中后代与祖先出现的相似中得到了进一步的验证。“优胜劣汰,适者生存”的法则在新物种的产生中得到了体现。因此,只要我用“突变”来替代“细微的偶然变异”,就是正确表达了多数生物学家的观点和理论,这其中自然也包括达尔文——除了“突变”的出现之外,他的学说中的其他方面无须修改[19] 。
3.定位,隐性和显性
我们再对突变的其他事实和概念进行有益的探索和讨论,虽然是形式化的评论,但也不需要直接说明它们是如何从实验数据中推导出来的。
一条染色体在其自身所在区域内发生突变是可以被我们直接观察到,并得以确认的。与这条染色体的变化相比较,同源染色体的对应位置上没有发生一点变化(图8给出了示意图,×表示突变的位置),这一点是十分重要的。通常在后代中有一半的突变体性状可以直接显现出来,另一半则保持正常没有变化,这刚好证明了在突变个体与非突变个体的杂交中一条染色体受到突变的影响,而另一条染色体没有变异。这和理论的预期一样,证明了是突变体发生减数分裂时两条染色体互相分离的结果——如图9所示。这个图是一个“谱系”,一对染色体表示了三代连续的个体。如果个体的突变体的两条染色体发生突变的话,那么其子女全会得到相同的遗传性——与他们的父本、母本都不相同。
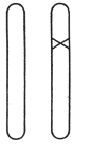
& & & & & 图8 杂合的突变体。 x标明突变的基因
然而,有一个无可争议的事实:这个领域内的实验十分复杂,并不是我们前面所说的那样简单。这是由于偶然性的突变发生的时候并非是显然的,而是潜移默化的。什么意思呢?
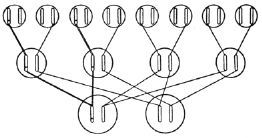
图9 突变的遗传。
交叉的连线表示染色体的传递,双线表示突变染色体的传递。
第三代的未说明来由的染色体来自图内未包括的其他第二代的配偶。
假定这些配偶不是亲戚,
也没有突变。
偶尔也有这样的情况出现,突变体里的两份“遗传密码本的拷贝”有些出入;至少发生突变的位置已经是两个不同的“密码”信息了。于是,有些人把原始的密码看做是“正统的”,把突变体的密码看做是“异端的”,这种认识是错误的;因为我们知道正常的突变也是从突变那里发展而来的。
在现实中,有两个版本,个体的“模式”在两者之中仿效一个。当然,这些版本可以是正常的,也可以是突变的。两者中被仿效的那个版本叫做显性,剩下的另一个版本叫做隐性。也就是说,根据模式的改变是否受到直接的突变,我们可以称之为显性突变或隐性突变。
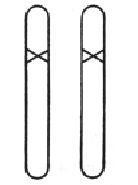
图10 纯合突变体,是从杂合突变体( 图8) 自体受精,或两个杂合突变体杂交产生的后代中的1/4的个体中获得。
隐性突变的概率有时甚至比显性突变要大,尽管它在突变开始发生的时候不明显,但它们的重要性是不言而喻的。只有两条染色体上都发生了隐性突变,才会影响到模式的改变(图10)。两个等同的隐性突变相互杂交或一个突变体自交时,就会产生这样的个体;在雌雄同体的植物那里这种情况是经常发生的,甚至是自发产生的。在这种情况中,可以观察到有隐性突变的模式在后代中约占四分之一。
4.介绍一些术语
解释一些专业术语,对于讲清问题还是有必要的。比如,在讲“密码的版本”——原始密码或者突变密码时,我实际上说的就是相同位置上的“等位基因”。如图8所示,密码的两个版本是不同的,相对于这个等位点来说个体就是杂合的。反之,如果这两个版本是完全相同的,如图10所示,这就是纯合的。于是我们可以看出,只有在纯合的情况下,模式的改变才是由隐性的等位基因所致。而在杂合个体或者纯全个体中的显性等位基因,都会产生相同的模式。
相对于白色来讲,只要有颜色,那几乎总是显性的。比如,豌豆的两个染色体,只有在其里面存有“白色的隐性等位基因”——也就是“白色纯合的”时候,它才会开出白色的花朵;它繁育出来的后代几乎都是一样的,都是开白色花朵的。但是,当一个“红色显性等位基因”(另一个基因是白色隐性的,个体是“杂合的”)出现时,它就会开出红色的花朵。如果有两个红色等位基因的话,那自然也是开红色的花朵了。由于杂合的红色可能会产生一些开白花的后代,而纯合的红色只能产生开红花的后代,所以上面说的后两种情况的差别只有在后代中才能显现出来。
有一个事实我们不容忽视,那就是外观上十分相似的两个个体,它们的遗传基因却有可能大不相同。所以,我们在探讨的时候需要明确区分。按照遗传学家们的惯常说法,它们的表现型虽然相同,但是遗传型却是不同的。
总结一下,我们可以用专业术语将前面几节的内容简短地概括为:
隐性等位基因只有在一种情况下才能影响遗传基因的表现型,那就是当这个遗传基因是纯合的时候。
这些专业性的说法,我们在后面还会遇到,在必要的时候我会向读者重申它的含义。
5.近亲繁殖的有害效应
只要发生隐性突变的个体是杂合的,自然选择就对它们毫无作用。一般而言,突变的发生往往都是有害的,只是由于它们是潜在的,因而自然选择一直没有消除它们。从某种程度上来说,长时间积累起来的有害突变是不会立刻造成明显的损害的。不幸的是,它们一定会把这种有害的基因传递给后代中的一半个体。有害基因的遗传规律对人、家禽、家畜或我们关心的其他物种的体质来说,都有着至关重要的应用。在图9中,假定一个男人(说具体些,比如我自己)在杂合的状态下,基因中携带有一个隐性的有害突变,正如前文所言,它不会在我身上明显地表现出来。如果这种突变没有出现在我的妻子身上的话,那么,我的一半数量的子女中(图9中的第二排)将会带有这种杂合的有害的隐性突变。如果我这样的子女与没有突变基因的配偶结婚(为了避免混淆,在图9中子女的配偶被省略了),那么,我的孙子、孙女中将会有1/4的人数将会受到以同样的方式来自遗传的有害突变的影响。
只有具有同样有害的隐性基因的个体互相杂交,后代的身上才会明显地表现出这种有害的危险。稍微回顾一下上文的内容,我们就可以知道这样杂交的结果会导致他们的后代中将有1/4的数量是纯合的,与之而来的则是危害性的表现。仅次于自体受精——这种情况只会出现在雌雄同株的植物上,与其相比,最大的危险是我的子女之间通婚。他们中的每一个人有没有这种隐性的潜在危害的机会是相等的,因此乱伦结合的话,其中有1/4是危险的,他们的后代中有1/4将表现出隐性基因的危害。因此,对于每一个乱伦生下来的孩子来说,其身上含有的危险因子的概率是1∶16[20] 。
同样的道理,我的两个“纯血缘的”孙儿、孙女结婚生下的后代的危险因子是1∶64。这种事情的发生似乎不太可能,但事实上这样的婚姻经常发生。可是,根据前面所分析过的理论依据,在祖代的配偶(“我和我的妻子”)中,一方已经带有一个可能的潜在危害的后果。事实上,他们两个人藏有的潜在的缺陷数目远远超过了一个。如果已经知道你身上有一个隐性的缺陷基因,那么就可以推断出,在你的8个堂、表兄妹之间也一定有一个是带有这种缺陷的。根据动植物的实验来看,隐性的危害基因中除了严重的、比较罕见的缺陷外——当然它们的数目是很少的,还有许多较小的缺陷。这样的话,这些大小缺陷的概率加起来就必然导致整个近亲繁殖的后代出现危害性状的概率大为增加,甚至使得他们严重衰退恶化。历史上,斯巴达人用非常残忍的方式消灭了失败者,但是我们现代人是不能用那样的方法的,我们必须严肃对待人类中的这类事情。在人类社会中,最适者生存的自然选择要少了许多,甚至转向了反面。
6.一般的和历史的陈述
在杂合的过程中,显性等位基因彻底掩盖了隐性等位基因,以至于我们很难发现会有什么样的效应出现。不过,这个令人惊奇的事实也有些许的例外。比方说,纯合的白色金鱼草与同样是纯合的深红色的金鱼草杂交,它们的所有直接后代都是中间型的颜色,即不是预期的深红色而是粉红色的。还有一个更为重要的例子,那就是血型。我们在这里不再对两个等位基因各自同时显示它们的影响。如果最后的探讨得出的结论是隐性可以分成不同的等级,而且用来检查“表现型”的实验的灵敏度在此过程中发挥关键作用,这完全在我的意料之中。
在这里有必要谈谈早期遗传学的历史,在遗传规律的发现方面,尤其是关于显性基因和隐性基因的重要区别,G.孟德尔(1822~1884)这位奥古斯丁教派的修道院长为其作出了巨大的贡献。对染色体与突变一无所知的孟德尔,在布隆(布尔诺)修道院的花园中,辛勤地播种着豌豆,来进行他所喜欢的实验。他栽种了几个不同品种的豌豆,让它们杂交,在它们成长的过程中注意观察各个后代的情况。实际上,在这个实验中,自然界中现成的突变体成了他实验的对象。1866年,在“布隆自然研究者协会”的会报上,他发表了这次实验的结果。那个年代,几乎没有一个人对他的实验和实验结果感兴趣。然而,人们没有想到的是,这个修道士的发现成为当代最有活力的学科,还成为了20世纪一门全新科学的纲领性原则。曾经发表过的论文也被人遗忘了,直到1900年,阿姆斯特丹的德佛里斯、维也纳的丘歇马克和柏林的柯灵斯三人同时各自独立地发现了这个结论,这个修道士的论文才被人们重新想起。
7.作为一种罕有事件,突变具有其必要性
到目前为止,我们一直都在关注有害突变。这种类型的突变相比较而言出现的频率会多一些,更重要的是会产生有害的性状;但我们也应该指出,有利突变的情形也是存在的。自发的突变是物种发展历史上的一个小插曲——以偶然的形式并且冒着存在有害因素而被自动消除的风险却不断作出“尝试”。由此我们可以得知,自然选择可以把突变作为自己的合适原料,但这种突变必须得像自然界中出现的那样罕见的事件。如果突变非常频繁的话,就会导致很多机会,比如说,在同一个体-内出现了十几个不同的突变,而其中有害突变远远超过有利突变,那么,物种不仅没有得到改良,反而停滞不前,甚至走向衰亡。基因的保守性是显而易见并且是十分必要的,这得益于它的高度的持久性。例如,一个大型制造厂的经营,为了创造一种更好的生产方法,大胆的革新即使没有得到确认,但却是必须的。其中的某些革新是促进生产力还是降低生产力,在一定的时间限度内只能采用一项革新而其他保持不变。
8.X射线诱发的突变
有关遗传学的一系列实验研究,我们可以一一回顾,因为这些将证明前面所谈到的那些重要的特性。
用X射线或g射线照射亲代,可以提高突变率,使得后代中的突变大于自然突变率所带来的突变。这种方式可以产生数量较多的突变,它与自然发生的相比,其实并没有多大的差异。因而,我们要想获得一种“自然”突变的话,不妨可以借用X射线的效用。
每一个特定的“转变”与正常的个体之间存在一个“X射线系数”,可以使正常的个体变成一个特殊的突变体,反之亦然。这是经过无数次的X射线突变实验,我们得出的结论。从生物学的角度看,这个系数表明了用单位剂量的X射线在子代出生之前照射亲体,由此造成的后代带有突变所占的百分比。
9.第一定律——突变是个单一性事件
诱发突变率的规律虽然很简单,但是却具有极大的启发性。第一定律是:
(1)突变频数的增加量随着射线剂量的增加而不断提升,因此对于这种比例关系,我们可以利用突变系数来表达。
简单的比例关系在我们眼中已经是很平淡的事情了,这便导致我们不经意间低估这一定律的后果。举个例子可以更好地理解这一点,比方说,商品的单价与商品的数量之间并不一定总是成比例的。通常买6个橘子的价格是一个水平,但是当你决定多买12个橘子时,他可能会以低于12个橘子的价钱卖给你。当橘子供不应求时,与刚才的情况恰好相反。于是,我们可以推断,假如一半剂量的辐射导致后代中千分之一发生突变,那么剩下的没有发生突变的后代是不受影响的——它们既不免于突变,也不倾向于突变。如果不是这样的话,另一半的辐射剂量就不会恰好再引起后代中的千分之一发生突变。因此,突变并不是由连续的小剂量辐射相互增强而引起的一种积累效应,上文的正比例变化规律便可以证明这一点。突变是单一性事件,并且它只是在辐射期间发生在一条染色体上。那么,哪一类事件属于这样的单一性事件呢?
10.第二定律——事件的局域性
(2)从软的X射线到相当硬的Y射线,如果广泛地改变射线的性质,只要给予的辐射剂量是相等的,那么突变系数就一直不会变化。
我们用伦琴单位来衡量射线剂量,换言之,射线剂量是由在照射下标准物质——温度为0℃,压力为1标准大气压(1.01×105帕)的空气——的单位体积内所能产生的离子总数来度量,并且这种标准物质是经过严格选择的。
由于平均相对原子质量与空气相同的元素组成了有机物的组织,所以选择空气作为标准物质。此外,选择空气作为标准物质也很方便。组织内电离作用或相关过程总量的下限[21] ,可以通过把空气中的电离数乘以二者的密度比得到。
我们可以从这个定律中知道,发生生殖细胞某个“临界”体积内的电离作用是造成突变的单一性事件。好奇的人们会问:这种临界体积有多大呢?要想回答这个问题,我们可以根据观察到的突变率估算出来。如果每立方厘米产生50000个离子的剂量,使得任意一个配子以某种特殊的方式在照射的区域里发生突变的概率是1∶1000,那么,我们就可以断定临界体积只有1/50000立方厘米的1/1000,换句话说就是只有五千万分之一立方厘米。这个推断出来的数字并不是确切的数字,只是为了说明一下问题。实际上,我们根据K.G.齐默尔、N.W.铁摩菲也夫和M.德尔勃吕克[22] 所写的一篇论文[23] ,可以得出实际估计的数字。他们写的这篇论文也是后面章节中要谈论的学说的主要来源。大约为10个平均原子距离的1个立方体,只包括大约1000个原子是他们在论文中得出的数据。换句话说,如果在距离染色体上某个特定的点不超过“十个原子距离”的范围内发生了一次电离(或激发)就有产生突变的一次机会。我们现在更详细地来讨论这一点。
铁摩菲也夫的报告隐含着一个十分重要的推论,我不得不在这里提一下,可能这与我们的研究没有什么实质性的关系。在现代生活中,人们或多或少地会接触到X射线的照射,这便会产生像X射线癌、烧伤、不育等这样一类比较直接性的危险。现在人们已用铅屏、铅围裙等作为防护来避免这些危险,尤其是对经常接触射线的护士和医生们,一定要为他们提供专业的保护。然而,即便这些对个人的直接危险,我们可以有效地防止,但是还存在着其他的间接危险——产生于生殖细胞内的有害突变,这也是我们在讨论近亲繁殖的不良后果时谈到的那种突变。说得严重一些的话,由于祖母是长期受X射线照射的护士,所以堂兄妹结婚的危害可能性会大大增加。当然,对于一个单独的个体来说,没有必要为此担心。但是对于包含个体的整个社会来说,这种潜在的有害突变是会慢慢地影响到人类的健康,因而需要我们人类充分而普遍的关注。